Введение
Спонтанные разряды с участием гиппокампа и парагиппокампальных структур являются отличительной чертой височной эпилепсии. Эпилептическая активность может далеко распространяться по синаптическим путям и приводить к сильной синхронизации между лимбической системою и другими областями мозга 1, 2. В наиболее широко используемых экспериментальных моделях височной эпилепсии спонтанные лимбические разряды развиваются после первоначального фармакологически или электрически индуцированного эпилептического статуса 3. Однако у неэпилептических крыс лимбические разряды могут возникать и при хроническом воздействии антагонистов эндоканнабиноидных CB1-рецепторов 4.
Эндоканнабиноидная система участвует в регуляции чувствительности мозга к эпилептической активации 5, 6. В опытах на крысах с использованием пилокарпиновой модели эпилептогенеза было показано, что введение агонистов CB1-рецепторов укорачивает ранние пост-статусные эпилептические разряды и уменьшает процент последующей смертности 7. Более того, в той же пилокарпиновой модели введение агонистов CB1-рецепторов полностью блокировало возникновение поздних спонтанных эпилептических разрядов 5. С другой стороны, антагонисты CB1-рецепторов снижают устойчивость мозга к судорогам 5. Установлено, что введение антагонистов CB1-рецепторов способствует распространению судорог от среднего мозга к лимбическим структурам у крыс, склонных к аудиогенным припадкам среднего мозга 8. Более того, здоровые крысы, которым в течение длительного времени делались инъекции антагониста CB1-рецепторов, становятся склонны к спонтанным разрядам 4. Именно такие спонтанные разряды, вызванные систематическим введением антагониста CB1-рецепторов, являются объектом данного исследования.
Эндоканнабиноиды синтезируются по требованию после активации постсинаптических нейронов и ретроградно подавляют высвобождение нейромедиаторов через пресинаптические метаботропные CB1-рецепторы. Таким образом, эндоканнабиноиды способствуют поддержанию физиологического уровня возбудимости нейронов и сохранению синаптических функций 9. Эпилептическое возбуждение быстро активирует этот защитный механизм против перевозбуждения 10. Эндоканнабиноидные CB1-рецепторы 11, 12 играют решающую роль в эндоканнабиноидзависимой защите от судорог, а локальное удаление этих рецепторов в гиппокампе сильно обостряет судороги 13.
Широко распространенные внелимбические области, включая как корковые, так и подкорковые структуры, все чаще признаются вовлеченными во вторичное обобщение лимбических разрядов у эпилептических крыс 14, 15 и в сложные парциальные припадки у пациентов с височной эпилепсией (ВЭ) 15, 16. Целью настоящего исследования был анализ направления функциональной связанности между неокортексом, гиппокампом, таламусом и средним мозгом во время иктального периода SLV326-индуцированных лимбических разрядов. Гиппокамп особенно подвержен эпилептическому возбуждению. Область среднего мозга содержит триггерные зоны для некоторых типов разрядов, в первую очередь для генерализованных тоник-клонических 17, 18, 19. Таламус является той областью мозга, которая ответственна за распространение эпилептических разрядов, также у пациентов с ВЭ взаимодействие таламуса с гиппокампом и корой изменяется по сравнению с нормальными контрольными испытуемыми 15, 20, 21.
Для получения надёжных результатов при анализе временных рядов важно использовать методы, способные решить ряд проблем: 1) нелинейность сигналов, 2) направленность связи, 3) причина сходства сигналов, включая прямые связи, общий источник, опосредованное воздействие через некоторую промежуточную структуру, или даже простое случайное совпадение колебаний 22. В данной работе для получения достоверных результатов использовался переменный во времени адаптированный метод причинности по Грейнджеру 23. Данная методика учитывает особенности экспериментальных данных: временные масштабы, нелинейность, эффективную размерность. Поэтому она имеет хорошее временное разрешение с оптимальной чувствительностью и специфичностью, как было показано 24, 25, будучи в основном нечувствительна к опосредованным связям 24.
Материалы и методы
Животные и записи локальных потенциалов мозга
Разряды, проанализированы в настоящем исследовании, являлись следствием хронических инъекций антагониста СВ1-рецепторов SLV326, как сообщалось ранее в 26. Поскольку методика выявления разрядов подробно описана в 26, здесь она будет изложена лишь вкратце.
Исследование проводилось в соответствии с руководящими принципами Европейского Сообщества по использованию экспериментальных животных и было одобрено этическим комитетом по изучению животных (RUDEC-2007-161).
Эксперименты проводились на 24 семимесячных крысах Crl:WI Wistar (Charles River Laboratories, Sulzfeld, Germany). С возраста 8-9 недель им ежедневно перорально через зонд вводили SLV326, 2-3 мг/кг, растворенного в полутвёрдом растворе. Разрешение на использование SLV326 было предоставлено Solvay Pharmaceuticals, Weesp, Нидерланды.
Всем 24 животным хирургически были имплантированы два триполярных ЭЭГ-электрода (Plastics One MS-332/2-A) под полной изофлурановой анестезией. Электроды были размещены в следующих структурах мозга (расстояния приведены в мм от брегмы: AP, L и H): лобная кора (FC): +2; -2; -1; гиппокамп (HP): -4.2; -3.6; -4.1; таламус (ТH): -2.6; -2.7; -7.3; и средний мозг (MB): -8.8; -1.7; -5.2. Заземляющий и референтный электроды размещались двусторонне над мозжечком, причём референтный электрод располагался сбоку ипсилатерально по отношению к регистрирующим электродам. Восстановление животных после операции занимало не менее двух недель.
Регистрация локальных потенциалов мозга (LFP) и видеорегистрация производились в течение 24 часов. Сигналы LFP усиливались, фильтровались от 1 до 100 Гц и оцифровывались с частотою дискретизации 512 Гц с использованием системы Windaq (DATAQ Instruments, Akron, OH).
В течение 24 часов 6 из 24 крыс продемонстрировали генерализованные лимбические разряды. В настоящем исследовании было проанализировано 30 разрядов, зарегистрированных у трёх крыс, так как для этих крыс все электроды демонстрировали соответствующий уровень сигнала в течение всей записи.
Применение переменного во времени адаптированного метода причинности по Грейнджеру к записям локальных потенциалов мозга
Для анализа связанности между четырьмя измеренными сигналами из разных областей мозга использовался переменный во времени адаптированный нелинейный метод причинности Грейнджера 23. Метод основан на построении двух предсказательных моделей: одномерной индивидуальной модели, которая предсказывает значения из ряда  на основе только значений этого ряда со средней ошибкой аппроксимации (1), и совместной модели, которая использует также данные из ряда
на основе только значений этого ряда со средней ошибкой аппроксимации (1), и совместной модели, которая использует также данные из ряда 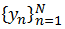 (её ошибка аппроксимации (2), как правило, меньше). Улучшение прогноза (3) является характеристикой связанности двух объектов и лежит в диапазоне от 0 до 1:
(её ошибка аппроксимации (2), как правило, меньше). Улучшение прогноза (3) является характеристикой связанности двух объектов и лежит в диапазоне от 0 до 1:
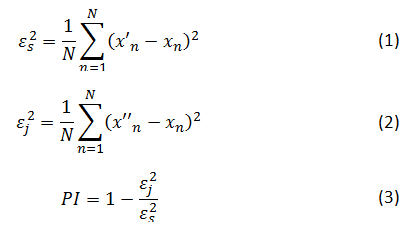
где xn — измеряемые значения, x'n соответствует значению, предсказанному с помощью индивидуальной модели, а x''n соответствует значению, предсказанному с помощью совместной модели.
Абсолютные значения улучшения прогноза PI обычно неинформативны, как это показали, например, Смирнов и Мохов 27. Однако увеличение или уменьшение PI имеет смысл при условии, что оператор эволюции рассматриваемых систем структурно не изменился (но его параметры могли измениться 24). Таким образом, изменения в силе связи могут быть обнаружены, если в скользящем временном окне использовать причинность по Грейнджеру 28. В настоящей работе использовалось временное окно длиной 1 с (512 точек данных) с перекрытием окон 0.9 с. Параметры метода, включая временные масштабы, были скорректированы в соответствии с рекомендациями и критериями, разработанными в 25, 29, 30.
Функция взаимной информации в скользящем окне
Функция взаимной информации (MI) вычислялась с помощью метода, предложенного в 31, для тех же пар каналов и в тех же условиях, что и причинность по Грейнджеру, включая длину и сдвиг временного окна, а также усреднение. Отметим, что MI — ненаправленная нелинейная мера похожести временных рядов. Это означает, что она не может показать направление воздействия или выявить причину изменений во взаимосвязи, как почти все простые меры, в том числе корреляция (линейная или нелинейная) или когерентность. Функция взаимной информации MI применяется к скалярным временным рядам. Современные работы 32, показывающие возможность значительно улучшить достоверность результатов с помощью векторных рядов с неоднородным вложением, основываются на слишком больших объёмах данных, что мешает применению этих методов в скользящих временных окнах. По этой же причине мы отказались от использования энтропии переноса 33, 34.
Статистический анализ оценок связанности
Для каждого разряда были рассчитаны индивидуальные зависимости улучшения прогноза от времени PI(t) и функции взаимной информации от времени MI(t). Анализируемые участки включали 10 секунд до начала разряда и 10 секунд после конца разряда. Соответственно, каждый рассматриваемый фрагмент ЭЭГ, содержащий лимбический разряд, был разделен на три части:
Начало: 10 с до и 10 с после начала разряда: преиктальный период и 10 с первой высокочастотной стадии;
Середина: 10 с до и 10 с после перехода от первой высокочастотной стадии ко второй низкочастотной стадии;
Конец: 10 с до и 10 с после окончания разряда: низкочастотная стадия и постиктальный период.
Полученные зависимости PI(t) и MI(t) были усреднены по всем разрядам, совмещая начало разряда, начало высокочастотной стадии и прекращение разряда. Затем для каждой усредненной зависимости PI(t) и MI(t) вычислялись фоновые уровни PIbg и MIbg, соответственно. Они считались как среднее значение на трёхсекундном временном интервале (фоновый период от 10 до 7 с до начала разряда). Используя средние фоновые уровни, вычислялись нормированные зависимости PI0 (t)=PI(t)-PIbg и MI0 (t)=MI(t)-MIbg. Значение PI0=0 (и MI0=0) соответствует фоновому уровню; положительные значения PI0 и MI0 соответствуют большему, чем в фоне, уровню связи, а отрицательные — меньшему.
Значения PI(t) и MI(t), полученные для различных разрядов в один и тот же момент времени, использовались в качестве выборки для выявления статистического отличия среднего значения от PIbg или MIbg соответственно, для чего был проведен одновыборочный t-тест. Поскольку этот тест проводился для каждой временной точки, была реализована Бонферрони-подобная коррекция для многократного тестирования: полученные p-значения умножались на число независимых временных интервалов, из которых рассчитывались PI и MI. Если после коррекции результирующее p-значение оказывалось меньше 0.05, то результаты рассматривались как существенно отличающиеся от фонового уровня и наносились на рис. 2, 3 цветом (синим, если PI<PIbg или MI<MIbg и красным, если PI>PIbg или MI>MIbg). В противном случае результаты были отмечены серым цветом как незначимые.
Результаты
Частотно-временной анализ
Все разряды начинались внезапно с резкого всплеска малой или средней амплитуды, который был обнаружен во всех зарегистрированных каналах одновременно. Общая продолжительность разрядов варьировала от 35.0 с до 115.3 с, а средняя продолжительность составляла 53.1 с. Можно выделить две основные стадии (рис. 1):
высокочастотная стадия, начинающаяся с частоты около 20 Гц, которая затем уменьшается до 15 Гц за первые 5 с;
низкочастотная стадия, в начале которой, основная частота колебаний резко падает до 2 Гц.
Высокочастотная стадия длилась от 23.3 с до 55.7 с при средней длительности 36.0 с. На этом этапе амплитуда колебаний возрастала. Вся стадия была очень нестационарна. Хорошо выраженные более высокие частоты (ещё выше основной частоты колебаний) можно наблюдать в разные моменты времени в течении этой стадии.
Низкочастотная стадия длилась от 5.7 с до 59.6 с при средней длительности 17.1 с. Высокочастотная динамика предыдущей стадии также присутствовала в первой части второй стадии, хотя была менее выражена. Низкочастотные колебания постепенно теряли амплитуду и внезапно прекращались. Подобные изменения считались маркером прекращения разряда.
Анализ направленного воздействия методом причинности по Грейнджеру
На рис. 2 и рис. 3 показаны результаты анализа связанности обоими методами: с помощью причинности по Грейнджеру и с помощью функции взаимной информации.
Инициация разряда
Примерно за 2 секунды до начала разряда во всех рассматриваемых парах каналов наблюдается падение связанности. Это падение продолжается в течение, по крайней мере, первых 10 секунд высокочастотной стадии (см. рис. 2, синие точки), за исключением пары гиппокамп — средний мозг, в которой взаимодействие восстанавливается до нормального уровня в начале разряда. Новое двунаправленное падение связанности в этой паре начинается примерно через 6 секунд после начала разряда.
Переход к низкочастотной стадии
Воздействие со стороны FC на HP остается ниже базового уровня в течение всего разряда (рис. 2c верхняя панель). При наличии некоторого кратковременного возрастания для остальных пар каналов связанность восстанавливается на протяжении высокочастотной стадии: однонаправленная от MB и HP на FC (рис. 2a и рис. 2b средние панели) и от TH на MB (верхняя панель рис. 3а), двунаправленная в паре HP–MB (рис. 3b верхняя и средняя панели).

Рисунок 1: Записи локальных потенциалов мозга во время лимбического разряда и их спектрограммы: (a, b) — из лобной коры, (c, d) — из таламуса, (e, f) — из гиппокампа, (g, h) — из среднего мозга. Три чёрные вертикальные линии отмечают: 1) начало разряда в момент времени 10 с, 2) границу между двумя стадиями в момент времени 38 с, и 3) конец разряда в момент времени 50 с.
Во время перехода от высокочастотной стадии к низкочастотной все связи примерно равны фоновому уровню за исключением пары FC–TH, в которой связанность уменьшается за 5 секунд до перехода (рис. 2b, верхняя и средняя панели), и кроме вышеупомянутой пары FC–HP, которая демонстрирует пониженную связанность в течение всего разряда (рис. 2с, верхняя панель).
В течение первых нескольких секунд низкочастотной стадии связанность ниже фонового уровня видна в направлении от FC и HP к MB (рис. 2a и 2e, верхние панели) и от TH к FC (рис. 2b, средняя панель). Воздействие со стороны FC и TH к HP также ниже фонового уровня. Эта пониженная связанность продолжается всю низкочастотную стадию (рис. 2c, верхняя панель, и рис. 3c, средняя панель).
Прекращение разряда
К концу разряда наблюдается переход к увеличению связанности — те же самые, что наблюдались в конце высокочастотной стадии: однонаправленное увеличение воздействие от MB и HP на FC (рис. 2a и рис. 2b средние панели), и от TH на MB (рис. 3а, верхняя панель), но двунаправленное увеличение взаимодействия в паре НР–MB, присутствующее в высокочастотной стадии, теперь становится статистически значимым только по направлению от HP к MB (рис. 3b, верхняя панель). Увеличение связанности от MB к HP отсутствует, в то время как это увеличение было очень выражено в конце высокочастотной стадии (рис. 3b, средняя панель). Кроме того, воздействие со стороны FC и TH на HP все ещё низкое (рис. 2c, верхняя панель, и рис. 3c, средняя панель).
Когда движущееся окно захватывает конец разряда, происходит резкое увеличение связанности во всех парах каналов, в которых таламус участвует в качестве управляемой структуры (рис. 2b, 3a и 3c). Однако такое увеличение упоминалось ранее как артефакт метода 24. В постиктальной стадии все связи, как правило, достаточно низкие по сравнению с фоновыми.

Рисунок 2: Зависимости среднего нормированного улучшения прогноза PI0 и функции взаимной информации MI0 от времени для пар каналов FC–MB, FC–Th, FC–Hp, посчитанные в скользящем окне длительностью 1 с. Красные и синие точки показывают значения, которые статистически значимо (p-значение < 0,05 с поправкой Бонферрони) отличаются (красные — в большую, синие — в меньшую сторону) от фонового уровня, серые точки — не отличающиеся от фонового уровня. Чёрные пунктирные вертикальные линии обозначают начало разряда, переход ко второй стадии и окончание, серая линия перед ними — отрезок скользящего временного окна. Фоновый уровень, первая (высокочастотная) стадия, вторая (низкочастотная) стадия и постиктальный период подписаны в верхней части каждого фрагмента рисунка.

Рисунок 3: Зависимости среднего нормированного улучшения прогноза PI0 и функции взаимной информации MI0 от времени для пар каналов Th–MB, Hp–MB, Hp–Th, посчитанные в скользящем окне длительностью 1 с. Красные и синие точки показывают значения, которые статистически значимо (p-значение < 0,05 с поправкой Бонферрони) отличаются (красные — в большую, синие — в меньшую сторону) от фонового уровня, серые точки — не отличающиеся от фонового уровня. Чёрные пунктирные вертикальные линии обозначают начало разряда, переход ко второй стадии и окончание, серая линия перед ними — отрезок скользящего временного окна. Фоновый уровень, первая (высокочастотная) стадия, вторая (низкочастотная) стадия и постиктальный период подписаны в верхней части каждого фрагмента рисунка.
Анализ взаимодействия с помощью расчёта функции взаимной информации
Результаты, полученные с помощью расчёта функции взаимной информации, отображаются в нижней панели каждого фрагмента рисунка. Кривые нормированной функции взаимной информации MI0 (t) сильно колеблются и большинство значений статистически незначимы. Функция взаимной информации значительно выше фонового уровня для пары FC–MB (рис. 2a), при этом для пар FC–HP и TH–HP она меньше фонового уровня (рис. 2c и рис. 3c). Для пары TH–MB за 5 с до окончания разряда ряд значений значимо превышают базовый уровень (рис. 3а).
Обсуждение
Полная схема комплексных изменений в сети представлена на рис. 4. Основные результаты могут быть сформулированы следующим образом:
В течение лимбических разрядов выделяются две стадии: высокочастотная (15-20 Гц), за которой следует низкочастотная (2 Гц).
Примерно за 2 секунды до начала разряда происходит падение связанности во всех рассматриваемых парах каналов. Это падение продолжается в течение, по крайней мере, первых 10 с высокочастотной стадии.
В течение всего разряда воздействие от лобной коры к гиппокампу остаётся сниженным. Для всех остальных пар каналов связь восстанавливается во время высокочастотной стадии с некоторым возрастанием во время переходных процессов.
В постиктальный период все взаимодействия достаточно низкие по сравнению с фоновыми.

Рисунок 4: Схемы значимых изменений связанности на разных стадиях разряда. Синие/красные стрелки показывают значительное снижение/увеличение улучшения прогноза PI0, значительные изменения в функции взаимной информации показаны рядом со стрелками знаками MI↑ (красный) и MI↓ (синий).
Обсуждение методов
Результаты анализа связанности сложных временных рядов всегда зависят от метода 22. Простые методы, такие как линейная корреляция и функция когерентности, могут быть легко вычислены с помощью стандартных наборов инструментов, таких как MATLAB или SciPy. Однако, как правило, считается, что эти методы не вполне адекватны по следующим причинам: 1) можно анализировать только линейные сходства, 2) направление связи не может быть обнаружено, 3) причина сходства остается неизвестной (однонаправленное движение, двунаправленное, общий источник, опосредованная связь через некоторую промежуточную структуру или просто случайное совпадение ритмов колебаний). Некоторые нелинейные меры, такие как взаимная информационная функция 31 (MI), коэффициент нелинейной корреляции 35, индекс фазовой синхронизации 36 преодолевают первую проблему, но не другие.
Ранее функция взаимной информации уже использовалась для изучения механизмов эпилепсии у животных 37 и человека 38. В качестве способа определения направления связи рассматривался сдвиг во времени между рядами, при котором мера демонстрировала максимум. Однако недавние исследования ясно показали, что неориентированные по своей природе нелинейные меры подобия, в том числе индексы фазовой синхронизации 39 и взаимная информация 40, небезопасны для определения направленности связи, так как очень часто результаты оказываются случайными и вводящими в заблуждение даже для относительно простых моделируемых рядов. В текущем исследовании была применена функция взаимной информации, рассчитанная с использованием наиболее выгодного подхода 31, но с весьма ограниченным успехом: многие результаты либо статистически незначимы, либо не могут быть связаны с результатами спектрального анализа или анализа связанности методом причинности по Грейнджеру.
Наиболее перспективными методами, позволяющими решить все три указанные проблемы, являются метод причинности по Грейнджеру 41, частная направленная когерентность 42, методы моделирования фазовой динамики 43 и энтропия переноса 33. Все эти методы являются более или менее параметрическими. Как правило, параметризация существенно снижает требования к объёмы данных. В принципе, информацию о направленной связи можно извлечь из относительно коротких рядов: 30 колебаний для фазовой динамики 43 и 4-8 колебаний для причинности по Грейнджеру 23. Однако риск получения ложноположительных (недостаточная специфичность) и ложноотрицательных (недостаточная чувствительность) результатов из-за неправильной параметризации также возрастает 25, 44, 45. До сих пор нет возможности получить абсолютную гарантию от ложных результатов, можно только снизить риски путем адаптации используемых методов к специфике исследуемых данных.
В данной работе был проведён попарный анализ всех пар каналов. Это небезопасно, так как прямые и косвенные связи не могут быть полностью разделены. Но классическая условная причинность по Грейнджеру 41, а также более продвинутые подходы 46, направленные на устранение избыточности исходных данных, не могут быть применены напрямую, так как требуют слишком много данных. Например, в 46 при использовании линейной причинности по Грейнджеру были задействованы 4000 точек данных (10 с записи, 400 Гц частота дискретизации), в то время как для изучения лимбических разрядов приходится работать с нелинейной причинностью (в этом случае требуется гораздо больше коэффициентов модели), иначе специфичность метода становится очень низкой, как это было показано в 41, 40 и 24. Кроме того, из-за нестационарности сигнала необходимо учитывать относительно короткие временные отрезки, не превышающие 1 с. Однако внутричерепные локальные потенциалы мозга гораздо менее избыточны, чем поверхностные ЭЭГ, особенно в том случае, если они были получены из различных и разнесённых пространственно структур мозга, как в настоящем исследовании.
Обсуждение изменения архитектуры связей во время разряда
Проведенный анализ основан на усреднении более 30 разрядов у трех крыс, получавших SLV-терапию (весь набор данных от 3 животных). В 47 обнаружена большая вариабельность результатов статистических моделей, построенных по данным ЭЭГ. Поэтому, в дополнение к уже описанным выше результатам результаты метода причинности по Грейнджеру были также усреднены отдельно по животным. Результаты, полученные на основе анализа данных одного животного, качественно выглядят также, как усреднённые по всем животным, но с меньшей значимостью. Поэтому выводы были сделаны на основе всего набора данных.
Снижение воздействия со стороны лобной коры на подкорковые структуры и увеличение воздействия со стороны подкорковых участков на неокортекс, описанные в ходе исследования генерализованных лимбических разрядов, согласуются с результатами визуализационных исследований как у эпилептических крыс 48, так и у пациентов с височной эпилепсией 15, 16. Эти исследования показали, что лимбические разряды сопровождаются активацией подкорковых структур и дезактивацией неокортекса. Выдвинута гипотеза, что аномально повышенная активность таламуса и верхнего ствола головного мозга препятствует нормальной активации коры и приводит к аномально пониженной функции лобно-теменной коры, что лежит в основе аномального двигательного поведения и нарушения сознания при сложных парциальных припадках 14, 15. Наши результаты показывают, что, несмотря на вовлечение лобной коры в экспрессию разрядов, она играет лишь пассивную роль, будучи управляемой подкорковыми структурами.
Основную активаторную роль играют гиппокамп и средний мозг. Обе структуры демонстрируют внутреннюю эпилептогенность и содержат триггерные зоны для определенных типов судорог: гиппокамп для лимбических судорог, средний мозг для рефлекторных аудиогенных судорог 18, 19. Эти две структуры посылают прямые и косвенные глутаматергические проекции в кору, таламус и друг другу. Гиппокамп и средний мозг могут являться активаторами разрядов, а их взаимное возбуждающее взаимодействие во время лимбических разрядов может представлять собой петлю положительной обратной связи, приводящий к разрядам.
Хотя центральная роль гиппокампа в возникновении припадков височной эпилепсии хорошо известна, роль среднего мозга в лимбических разрядах оставляет вопросы. Тем не менее, в работе показано участие среднего мозга в механизмах вторичных генерализованных лимбических разрядов 49 и первичных генерализованных тоник-клонических разрядов 17. Ранее было показано, что хроническое введение других антагонистов CB1-рецепторов (римонабант) значительно облегчает распространение судорог от среднего мозга к коре во время аудиогенного киндлинга 8. Интересно исследовать, является ли активное участие среднего мозга в лимбических разрядах обычным явлением, или оно наблюдается только при лимбических разрядах, индуцированных антагонистами каннабиноидов.
Заключение
Результаты нашего исследования показывают, что кортико-таламическая петля, лежащая в основе поддержания абсансных разрядов 50, 51, демонстрирует падение в связанности в течение всего генерализованного лимбического разряда, что соответствует снижению таламо-кортикальной функциональной связности у пациентов с ВЭ 20. С другой стороны, во время лимбических разрядов лобная кора воздействует на гиппокамп, который активно вовлекается в механизмы генерации в отличие от абсансных разрядов, для которых наблюдалось падение связанности в кортико-гиппокампальных и кортико-таламических парах 52.
Благодарности
Данная работа была выполнена при поддержке Стипендии Президента РФ для молодых учёных СП-3605.2018.4
- Bertram, E. H.The functional anatomy of spontaneous seizures in a rat model of chronic limbic epilepsy. Epilepsia 1997; 38(1): 95-105 (https://doi.org/10.1111/j.1528-1157.1997.tb01083.x).
- Spenser, S. S.Neural networks in human epilepsy: evidence of and implications for treatment. Epilepsia 2002; 43(3): 219-227 (https://doi.org/10.1046/j.1528-1157.2002.26901.x).
- Curia, G., Longo, D., Biagini, G., Jones, R. S. G., Avoli, M.The pilocarpine model of temporal lobe epilepsy. J Neurosci Methods 2008; 172(2): 143-157 (https://doi.org/10.1016/j.jneumeth.2008.04.019).
- van Rijn, C. M., Perescis, M. F. J., Vinogradova, L., van Luijtelaar, G.The endocannabinoid system protects against cryptogenic seizures. Pharmacological Reports 2011; 63(1): 165-168 (https://doi.org/10.1016/S1734-1140(11)70411-X).
- Wallace, M. J., Blair, R. E., Falenski, K. W., Martin, B. R., DeLorenzo, R. J.The endogenous cannabinoid system regulates seizure frequency and duration in a model of temporal lobe epilepsy. J Pharmacol. Exp. Ther. 2003; 307(1): 129-137 (https://doi.org/10.1124/jpet.103.051920).
- Alger, B. E.Endocannabinoids and their implication for epilepsy. Epilepsy Curr. 2004; 4(5): 169-173 (https://doi.org/10.1111/j.1535-7597.2004.04501.x).
- Suleymanova, E. M., Shangaraeva, V. A., van Rijn, C. M., Vinogradova, L. V.The cannabinoid receptor agonist WIN55.212 reduces consequences of status epilepticus in rats. Neuroscience 2016; 334(): 191-200 (https://doi.org/10.1016/j.neuroscience.2016.08.004).
- Vinogradova, L. V., Shatskova, A. B., van Rijn, C. M.Pro-epileptic effects of the endocannabionoid receptor antagonist SR141716 in a model of audiogenic epilepsy. Epilepsy Research 2011; 96(3): 250-256 (https://doi.org/10.1016/j.eplepsyres.2011.06.007).
- Katona, I. & Freund, T. F.Multiple functions of endocannabinoid signaling in the brain. Ann. Rev. Neurosci. 2012; 35(): 529-558 (https://doi.org/10.1146/annurev-neuro-062111-150420).
- Lutz, B.On-demand activation of the endocannabinoid system in the control of neuronal excitability and epileptiform seizures. Biochem. Pharmacol. 2004; 68(9): 1691-1698 (https://doi.org/10.1016/j.bcp.2004.07.007).
- Fernandez-Ruiz, J. & Gonzales, S., Cannabinoid control of motor function at the basal ganglia In: Handbook of Experimental Pharmacology: Cannabinoids. Springer; 2005: 479-507 (https://doi.org/10.1007/3-540-26573-2_16).
- M Herkenham, A B Lynn, M D Little, M R Johnson, L S Melvin, B R de Costa, and K C Rice Cannabinoid receptor localization in brain. Proceedings of the National Academy of Sciences 1990; 87(5): 1932-1936 (http://www.pnas.org/content/87/5/1932).
- Krisztina Monory, Federico Massa, Michaela Egertová, Matthias Eder, Heike Blaudzun, Ruth The Endocannabinoid System Controls Key Epileptogenic Circuits in the Hippocampus. Neuron 2006; 51(4): 455–466 (https://doi.org/10.1016/j.neuron.2006.07.006).
- Norden, A. D. & Blumenfeld H.The role of subcortical structures in human epilepsy. Epilepsy and Behavior 2002; 3(3): 219–231 (https://doi.org/10.1016/S1525-5050(02)00029-X).
- Blumenfeld, H., Varghese, G. I., Purcaro, M. J., Motelow, J. E., Enev, M., McNally, K. A., Levin, A. R., Hirsch, L. J., Tikofsky, R., Zubal, I. G., Paige, A. L., Spencer, S. S.Cortical and subcortical networks in human secondarily generalized tonic–clonic seizures. Brain 2009; 132(4): 999-1012 (https://doi.org/10.1093/brain/awp028).
- Haneef, Z., Lenartowicz, A., Yeh, H. J., Levin, H. S., Engel, J. Jr., Stern, J. M.Functional connectivity of hippocampal networks in temporal lobe epilepsy. Epilepsia 2014; 55(1): 137–145 (https://doi.org/10.1111/epi.12476).
- Browning, R. A. & Nelson, D. K.Modification of electroshock and pentylenetetrazol seizure patterns in rats after precollicular transections. Experimental Neurology 1986; 93(3): 546-556 (https://doi.org/10.1016/0014-4886(86)90174-3).
- Gale, K.Subcortical structures and pathways involved in convulsive seizure generation. J Clin Neurophysiol 1992; 9(2): 264-277 (http://europepmc.org/abstract/MED/1350593).
- McCown, T. J., Greenwood, R. S., Breese, G. R.Inferior Collicular Interactions with Limbic Seizure Activity. Epilepsia 1987; 28(3): 234–241 (https://doi.org/10.1111/j.1528-1157.1987.tb04213.x).
- He, X., Doucet, G. E., Sperling, M., Sharan, A., Tracy, J. I.Reduced thalamocortical functional connectivity in temporal lobe epilepsy. Epilepsia 2015; 56(10): 1571–1579 (https://doi.org/10.1111/epi.13085).
- Dinkelacker, V., Valabregue, R., Thivard, L., Lehéricy, S., Baulac, M., Samson, S., Dupont, S.Hippocampal-thalamic wiring in medial temporal lobe epilepsy: Enhanced connectivity per hippocampal voxel. Epilepsia 2015; 56(8): 1217–1226 (https://doi.org/10.1111/epi.13051).
- Gourevitch, B., Le Bouquin-Jeannes, R., Faucon, G.Linear and nonlinear causality between signals: methods, examples and neurophysiological applications. Biological Cybernetics 2006; 95(4): 349-369 (https://doi.org/10.1007/s00422-006-0098-0).
- Sysoeva, M. V., Sitnikova, E., Sysoev, I. V., Bezruchko, B. P., van Luijtelaar, G.Application of adaptive nonlinear Granger causality: Disclosing network changes before and after absence seizure onset in a genetic rat model. J Neurosci. Methods 2014; 226(): 33-41 (https://doi.org/10.1016/j.jneumeth.2014.01.028).
- Sysoev, I. V. & Sysoeva, M. V.Detecting changes in coupling with Granger causality method from time series with fast transient processes. Physica D: Nonlinear Phenomena 2015; 309(https://doi.org/10.1016/j.physd.2015.07.005): 9-19 ().
- Kornilov, M. V., Medvedeva, T. M., Bezruchko, B. P., Sysoev, I. V.Choosing the optimal model parameters for Granger causality in application to time series with main timescale. Chaos, Solitons & Fractals 2016; 82(): 11-21 (https://doi.org/10.1016/j.chaos.2015.10.027).
- Perescis, M. F. J., de Bruin, N., Heijink, L., Kruse, C., Vinogradova, L., Lüttjohann, A., van Luijtelaar, G., van Rijn, C. M.Cannabinoid antagonist SLV326 induces convulsive seizures and changes in the interictal EEG in rats. PLoS ONE 2017; 12(2): e0165363 ().
- Smirnov, D. A. & Mokhov, I. I.From Granger causality to long-term causality: Application to climatic data. Phys. Rev. E 2009; 80(): 016208 (https://doi.org/10.1103/PhysRevE.80.016208).
- Hesse, W., Möller, E., Arnold, M., Schack, B.The use of time-variant EEG Granger causality for inspecting directed interdependencies of neural assemblies. J Neurosci. Methods 2003; 124(1): 27-44 (https://doi.org/10.1016/S0165-0270(02)00366-7).
- Sysoeva M. V., Dikanev T.V., Sysoev I. V.Selecting time scales for empirical model construction. Izvestiya VUZ, Applied Nonlinear Dynamics 2012; 20(2): 54-62 (https://doi.org/10.18500/0869-6632-2012-20-2-54-62).
- Kornilov M. V., Sysoev I. V.Investigating nonlinear Granger causality method efficiency at strong synchronization of systems. Izvestiya VUZ, Applied Nonlinear Dynamics 2014; 22(4): 66-76 (https://doi.org/10.18500/0869-6632-2014-22-4-66-76).
- Kraskov, A., Stögbauer, H., Grassberger, P.Estimating mutual information. Phys. Rev. E 2004; 69(): 066138 (https://doi.org/10.1103/PhysRevE.69.066138).
- Kugiumtzis DDirect-coupling information measure from nonuniform embedding. Phys. Rev. E 2013; 87(): 062918 (https://doi.org/10.1103/PhysRevE.87.062918).
- Schreiber, T.Measuring Information Transfer. Phys. Rev. Lett. 2000; 85(): 461 (https://doi.org/10.1103/PhysRevLett.85.461).
- Montalto A, Faes L, MarinazzoDMuTE: A MATLAB Toolbox to Compare Established and Novel Estimators of the Multivariate Transfer Entropy. PLoS One 2014; 9(10): e109462 ( https://doi.org/10.1371/journal.pone.0109462).
- Pijn, J. P. M., Vijn, P. C. M., Lopes da Silva, F. H., Van Ende Boas, W., Blanes, W.Localization of epileptogenic foci using a new signal analytical approach. Neurophysiol. Clin. 1990; 20(): 1-11 (https://doi.org/ 10.1016/S0987-7053(05)80165-0 (PMID:2348808)).
- Allefeld, C. & Kurths, J.Testing for phase synchronization. Int. J. Bif. Chaos 2004; 14(2): 405-416 (https://doi.org/10.1142/S021812740400951X2).
- Mars NJI, Lopes da Silve, FHPropagation of seizure activity in kindled dogs. Electroencephalography and clinical neurophysiology 1983 ; 56(2): 194-209 (https://doi.org/10.1016/0013-4694(83)90074-3).
- Mars NJI, Thompson PM, Wilkus RJSpread of epileptic seizure activity in humans. Epilepsia 1985; 26(1): 85-94 (https://doi.org/10.1111/j.1528-1157.1985.tb05192.x).
- Vakorin VA, Mišić B, Krakovska O, Bezgin G, McIntosh RConfounding Effects of Phase Delays on Causality Estimation. PLoS One 2013; 8(1): e53588 ( https://doi.org/10.1371/journal.pone.0053588).
- Smirnov DAQuantifying causal couplings via dynamical effects: A unifying perspective. Phys. Rev. E 2014; 90(): 062921 (https://doi.org/10.1103/PhysRevE.90.062921).
- Chen YH, Rangarajan G, Feng JF, Ding MZ Analyzing multiple nonlinear time series with extended Granger causality. Phys. Lett. A 2004; 324(1): 26-35 (https://doi.org/10.1016/j.physleta.2004.02.032).
- Schelter, B., Timmer, J., Eichler, M.Assessing the strength of directed influences among neural signals using renormalized partial directed coherence. J. Neurosci. Methods 2009; 179(1): 121-130 (https://doi.org/10.1016/j.jneumeth.2009.01.006 (PMID: 19428518)).
- Smirnov, D. A. & Bezruchko, B. P.Estimation of interaction strength and direction from short and noisy time series. Phys. Rev. E 2003; 68(): 046209 (https://doi.org/10.1103/PhysRevE.68.046209).
- Smirnov, D. A. & Bezruchko, B. P.Spurious causalities due to low temporal resolution: Towards detection of bidirectional coupling from time series. Europhys. Lett. 2012; 100(1): 10005 (http://stacks.iop.org/0295-5075/100/i=1/a=10005).
- Smirnov, D. A.Spurious causalities with transfer entropy. Phys. Rev. E 2013; 87(): 042917 (https://doi.org/10.1103/PhysRevE.87.042917).
- Stramaglia S, Cortes JM, Marinazzo DSynergy and redundancy in the Granger causal analysis of dynamical networks. New Journal of Physics 2014; 16(10): 105003 (http://stacks.iop.org/1367-2630/16/i=10/a=105003).
- Papadopoulou M, Cooray G, Rosch R, Moran R, Marinazzo D, Friston KDynamic causal modelling of seizure activity in a rat model. Neuroimage 2017; 146(): 518-532 (https://doi.org/10.1016/j.neuroimage.2016.08.062 (PMID: 27639356)).
- Englot, D. J., Modi, B., Mishra, A. M., DeSalvo, M., Hyder, F., Blumenfeld, H.Cortical Deactivation Induced by Subcortical Network Dysfunction in Limbic Seizures. Journal of Neuroscience 2009; 29(41): 13006-13018 (https://doi.org/10.1523/JNEUROSCI.3846-09.2009 ).
- Chiba, S. & Wada, J. A.Amygdala kindling in rats with brainstem bisection. Brain Research 1995; 682(1-2): 50–54 (https://doi.org/10.1016/0006-8993(95)00315-H).
- Meeren, H. K., Veening, J. G., Möderscheim, T. A., Coenen, A. M., van Luijtelaar, G.Thalamic lesions in a genetic rat model of absence epilepsy: dissociation between spike-wave discharges and sleep spindles. Experimental Neurology 2009; 217(1): 25-37 (https://doi.org/10.1016/j.expneurol.2009.01.009 (PMID: 19416679)).
- Stefan, H. & Lopes da Silva, F. H.Epileptic neuronal networks: methods of identification and clinical relevance. Frontiers in Neurology 2013; 4(): Article 8 (https://doi.org/10.3389/fneur.2013.00008).
- Sysoeva, M. V., Vinogradova L. V., Kuznetsova, G. D., Sysoev, I. V., van Rijn, C. M.Changes in corticocortical and corticohippocampal network during absence seizures in WAG/Rij rats revealed with time varying Granger causality. Epilepsy and Behavior 2016; 64(Part A): 44–50 (https://doi.org/10.1016/j.yebeh.2016.08.009 (PMID: 27728902)).
Поступила в редакцию 09 января 2019 г., Принята в печать 03 февраля 2019 г.
